Модель ситуации типа «хищник-жертва. Курсовая работа: Качественное исследование модели хищник-жертва Математическая модель консервативная хищник жертва
Федеральное агентство по образованию
Государственное образовательное учреждение
высшего профессионального образования
«Ижевский государственный технический университет»
Факультет «Прикладная математика»
Кафедра «Математическое моделирование процессов и технологий»
Курсовая работа
по дисциплине «Дифференциальные уравнения»
Тема: «Качественное исследование модели хищник-жертва»
Ижевск 2010
ВВЕДЕНИЕ
1. ПАРАМЕТРЫ И ОСНОВНОЕ УРАВНЕНИЕ МОДЕЛИ «ХИЩНИК-ЖЕРТВА»
2.2 Обобщенные модели Вольтера типа «хищник-жертва».
3. ПРАКТИЧЕСКОЕ ПРИМЕНЕНИЕ МОДЕЛИ «ХИЩНИК-ЖЕРТВА»
ЗАКЛЮЧЕНИЕ
СПИСОК ЛИТЕРАТУРЫ
ВВЕДЕНИЕ
В настоящее время задачи экологии имеют первостепенное значение. Важным этапом решения этих задач является разработка математических моделей экологических систем.
Одной из основных задач экологии па современном этапе является изучение структуры и функционирования природных систем, поиск общих закономерностей. Большое влияние на экологию оказала математика, способствующая становлению математической экологии, особенно такие её разделы, как теория дифференциальных уравнений, теория устойчивости и теория оптимального управления.
Одной из первых работ в области математической экологии была работа А.Д. Лотки (1880 - 1949), который первый описал взаимодействие различных популяций, связанных отношениями хищник - жертва. Большой вклад в исследование модели хищник -жертва внесли В. Вольтерра (1860 - 1940), В.А. Костицин (1883-1963) В настоящее время уравнения описывающие взаимодействие популяций, называются уравнениями Лотки - Вольтерра.
Уравнения Лотки - Вольтерра описывают динамику средних величин - численности популяции. В настоящее время на их основе построены более общие модели взаимодействия популяций, описываемые интегро-дифференциальными уравнениями, исследуются управляемые модели хищник - жертва.
Одной из важных проблем математической экологии является проблема устойчивости экосистем, управления этими системами. Управление может осуществляться с целью перевода системы из одного устойчивого состояния в другое, с целью её использования или восстановления.
1. ПАРАМЕТРЫ И ОСНОВНОЕ УРАВНЕНИЕ МОДЕЛИ ХИЩНИК-ЖЕРТВА
Попытки математического моделирования динамики как отдельных биологических популяций, так и сообществ, включающих взаимодействующие популяции различных видов, предпринимались давно. Одна из первых моделей роста изолированной популяции (2.1) была предложена еще в 1798 г. Томасом Мальтусом:
, (1.1)Данная модель задается следующими параметрами:
N - численность популяции;
- разность между коэффициентами рождаемости и смертности.Интегрируя это уравнение получаем:
, (1.2)где N(0) – численность популяции в момент t = 0. Очевидно, что модель Мальтуса при
> 0 дает бесконечный рост численности, что никогда не наблюдается в природных популяциях, где ресурсы, обеспечивающие этот рост, всегда ограничены. Изменения численности популяций растительного и животного мира нельзя описывать простым законом Мальтуса, на динамику роста влияют многие взаимосвязанные причины – в частности, размножение каждого вида саморегулируется и видоизменяется так, чтобы этот вид сохранялся в процессе эволюции.Математическим описанием этих закономерностей занимается математическая экология – наука об отношениях растительных и животных организмов и образуемых ими сообществ между собой и с окружающей средой.
Наиболее серьезное исследование моделей биологических сообществ, включающих в себя несколько популяций различных видов, было проведено итальянским математиком Вито Вольтерра:
, - численность популяции; - коэффициенты естественного прироста (или смертности) популяции; - коэффициенты межвидового взаимодействия. В зависимости от выбора коэффициентов модель описывает либо борьбу видов за общий ресурс, либо взаимодействие типа хищник - жертва, когда один вид является пищей для другого. Если в работах других авторов основное внимание уделялось построению различных моделей, то В. Вольтерра провел глубокое исследование построенных моделей биологических сообществ. Именно с книги В. Вольтерра, по мнению многих ученых, началась современная математическая экология.2. КАЧЕСТВЕННОЕ ИССЛЕДОВАНИЕ ЭЛЕМЕНТАРНОЙ МОДЕЛИ «ХИЩНИК-ЖЕРТВА»
2.1 Модель трофического взаимодействия по типу «хищник-жертва»
Рассмотрим модель трофического взаимодействия по типу «хищник-жертва», построенную В. Вольтерром. Пусть имеется система, состоящая из двух видов, из которых один поедает другой.
Рассмотрим случай, когда один из видов является хищником, а другой - жертвой, и будем считать, что хищник питается только жертвой. Примем следующую простую гипотезу:
- коэффициент прироста жертвы; - коэффициент прироста хищника; - численность популяции жертвы; - численность популяции хищника; - коэффициент естественного прироста жертвы; - скорость потребления жертвы хищником; - коэффициент смертности хищника в отсутствие жертвы; - коэффициент «переработки» хищником биомассы жертвы в собственную биомассу.Тогда динамика численности популяций в системе хищник - жертва будет описываться системой дифференциальных уравнений (2.1):
(2.1)где все коэффициенты положительные и постоянные.
Модель имеет равновесное решение (2.2):
(2.2)По модели (2.1) доля хищников в общей массе животных выражается формулой (2.3):
(2.3)Анализ устойчивости состояния равновесия по отношению к малым возмущениям показал, что особая точка (2.2) является «нейтрально» устойчивой (типа «центр»), т. е. любые отклонения от равновесия не затухают, но переводят систему в колебательный режим с амплитудой, зависящей от величины возмущения. Траектории системы на фазовой плоскости
имеют вид замкнутых кривых, расположенных на различных расстояниях от точки равновесия (рис. 1).Рис. 1 – Фазовый «портрет» классической вольтерровой системы «хищник-жертва»
Разделив первое уравнение системы (2.1) на второе, получим дифференциальное уравнение (2.4) для кривой на фазовой плоскости
. (2.4)Интегрируя данное уравнение получим:
(2.5) - постоянная интегрирования, гдеНесложно показать, что движение точки по фазовой плоскости будет происходить только в одну сторону. Для этого удобно сделать замену функций
и , перенеся начало координат на плоскости в стационарную точку (2.2) и введя затем полярные координаты: (2.6)В таком случае, подставив значения системы (2.6) в систему (2.1), будем иметь.
В модели Колмогорова сделано одно существенное допущение: поскольку предполагается, что при то это означает существование в популяции жертв механизмов, регулирующих их численность даже в отсутствие хищников.
К сожалению, подобная формулировка модели не позволяет ответить на вопрос, вокруг которого в последнее время ведется много споров и о котором мы уже упоминали в начале главы: каким образом популяция хищников может оказывать регулирующее воздействие на популяцию жертв с тем, чтобы вся система была устойчива? Поэтому мы вернемся к модели (2.1), в которой механизмы саморегуляции (например, регулирование при помощи внутривидовой конкуренции) в популяции жертв (впрочем, как и в популяции хищников) отсутствуют; поэтому единственным механизмом регуляции численностей, входящих в сообщество видов, являются трофические взаимоотношения между хищниками и жертвами.
Здесь (так что, в отличие от предыдущей модели, Естественно, что решения (2.1) зависят от конкретного вида трофической функции которая, в свою очередь, определяется характером хищничества, т. е. трофической стратегией хищника и защитной стратегией жертвы. Общими для всех этих функций (см. рис. И) являются следующие свойства:
Система (2.1) имеет одну нетривиальную стационарную точку, координаты которой определяются из уравнений
![]()
при естественном ограничении .
Существует еще одна стационарная точка (0, 0), соответствующая тривиальному равновесию. Нетрудно показать, что эта точка - седло, а сепаратрисами являются координатные оси.
Характеристическое уравнение для точки имеет вид
![]()
Очевидно, что для классической вольтерровской модели .
Поэтому величину f можно рассматривать как меру отклонения рассматриваемой модели от вольтерровской.
![]()
стационарная точка - фокус, и в системе появляются колебания; при выполнении противоположного неравенства - узел, и колебания в системе отсутствуют. Устойчивость этого состояния равновесия определяется условием
т. е. существенным образом зависит от вида трофической функции хищника.
Условие (5.5) можно интерпретировать следующим образом: для устойчивости нетривиального равновесия системы хищник - жертва (и тем самым, для существования этой системы) достаточно, чтобы в окрестности этого состояния относительная доля потребляемых хищником жертв возрастала с возрастанием численности жертв. Действительно, доля жертв (от общей их численности), потребляемых хищником, описывается дифференцируемой функцией условие возрастания которой (положительность производной) выглядит как
![]()
Последнее условие, взятое в точке есть не что иное, как условие (5.5) устойчивости равновесия. При непрерывности оно должно выполняться и в некоторой окрестности точки Таким образом, если численности жертв в этой окрестности, то
Пусть теперь трофическая функция V имеет вид, изображенный на рис. 11, а (характерный для беспозвоночных). Можно показать, что для всех конечных значений (поскольку выпукла вверх)
т. е. при любых величинах стационарной численности жертв неравенство (5.5) не выполняется.
Это означает, что в системе с таким видом трофической функции не существует устойчивого нетривиального равновесия. Возможны несколько исходов: либо численности как жертвы, так и хищника неограниченно возрастают, либо (при прохождении траектории вблизи одной из координатных осей) в силу случайных причин численность жертвы или численность хищника станет равной нулю. При гибели жертвы через некоторое время погибнет и хищник, если же сначала погибает хищник, то численность жертвы начнет экспоненциально возрастать. Третий вариант - возникновение устойчивого предельного цикла - невозможен, что легко доказывается.
В самом деле, выражение

в положительном квадранте всегда положительно, если только имеет вид, изображенный на рис. 11, а. Тогда, согласно критерию Дюлака в этой области отсутствуют замкнутые траектории и устойчивого предельного цикла существовать не может.
Итак, можно сделать вывод: если трофическая функция имеет вид, изображенный на рис. 11, а, то хищник не может быть регулятором, обеспечивающим стабильность популяции жертвы и тем самым устойчивость всей системы в целом. Система может быть устойчива только в том случае, когда в популяции жертвы существуют собственные внутренние регулирующие механизмы, например, внутривидовая конкуренция или эпизоотии. Этот вариант регулирования был уже рассмотрен в §§ 3, 4.
Ранее отмечалось, что такой тип трофической функции характерен для хищников-насекомых, «жертвами» которых также обычно являются насекомые. С другой стороны, наблюдения за динамикой многих природных сообществ типа «хищник - жертва», включающих в себя виды насекомых, показывают, что для них характерны колебания очень большой амплитуды и весьма специфического вида.
Обычно после более или менее постепенного роста численности (который может происходить либо монотонно, либо в виде колебаний с возрастающей амплитудой) происходит ее резкое падение (рис. 14), и затем картина повторяется. По-видимому, такой характер динамики численностей видов насекомых может быть объяснен неустойчивостью этой системы при малых и средних значениях численности и действием мощных внутрипопуляционных регуляторов численности при больших ее значениях.

Рис. 14. Динамика численности популяции австралийской листоблошки Cardiaspina albitextura, кормящейся на эвкалиптах. (Из статьи: Clark L. R. The population dynamics of Cardiaspina albitextura.-Austr. J. Zool., 1964, 12, № 3, p. 362-380.)
Если же в систему «хищник - жертва» входят виды, способные к достаточно сложному поведению (например, хищники способны к обучению или жертвы способны находить убежище), то в такой системе возможно существование устойчивого нетривиального равновесия. Доказывается это утверждение весьма просто.
В самом деле, трофическая функция тогда должна иметь вид, изображенной на рис. 11, в. Точка на этом графике - точка касания прямой, проведенной из начала координат, графика трофической функции Очевидно, что в этой точке функция имеет максимум. Также легко показывается, что для всех выполняется условие (5.5). Следовательно, нетривиальное равновесие, в котором численность жертв меньше будет асимптотически устойчиво
Однако мы ничего не можем сказать о том, сколь велика область устойчивости этого равновесия. Например, если существует неустойчивый предельный цикл, то эта область должна лежать внутри цикла. Или другой вариант: нетривиальное равновесие (5.2) неустойчиво, но существует устойчивый предельный цикл; в этом случае также можно говорить о стабильности системы хищник - жертва. Поскольку выражение (5.7) при выборе трофической функции типа рис. 11, в может менять знак при изменении при , то критерий Дюлака здесь не работает и вопрос о существовании предельных циклов остается открытым.

Здесь в отличие от (3.2.1) знаки (-012) и (+a2i) разные. Как и в случае конкуренции (система уравнений (2.2.1)), начало координат (1) для этой системы является особой точкой типа «неустойчивый узел». Три других возможных стационарных состояния:

Биологический смысл требует положительности величин Х у х 2. Для выражения (3.3.4) это означает, что
В случае, если коэффициент внутривидовой конкуренции хищников а ,22 = 0, условие (3.3.5) приводит к условию ai2
Возможные типы фазовых портретов для системы уравнений (3.3.1) представлены на рис. 3.2 a-в. Изоклины горизонтальных касательных представляют собой прямые
а изоклины вертикальных касательных - прямые
Из рис. 3.2 видно следующее. Система хищник -жертва (3.3.1) может иметь устойчивое положение равновесия, в котором популяция жертв полностью вымерла (х = 0) и остались только хищники (точка 2 на рис. 3.26). Очевидно, такая ситуация может реализоваться лишь в случае, если кроме рассматриваемого вида жертв х хищник Х 2 имеет дополнительные источники питания. Этот факт в модели отражается положительным членом в правой части уравнения для хз. Особые точки (1) и (3) (рис. 3.26) являются неустойчивыми. Вторая возможность - устойчивое стационарное состояние, в котором популяция хищников полностью вымерла и остались одни жертвы - устойчивая точка (3) (рис. 3.2а). Здесь особая точка (1) - также неустойчивый узел.
Наконец, третья возможность - устойчивое сосуществование популяций хищника и жертвы (рис. 3.2 в), стационарные численности которых выражаются формулами (3.3.4). Рассмотрим этот случай подробнее.
Предположим равенство нулю коэффициентов внутривидовой конкуренции (аи = 0, i = 1, 2). Предположим также, что хищники питаются только жертвами вида х и в отсутствие их вымирают со скоростью С2 (в (3.3.5) С2
Проведем подробное исследование этой модели, воспользовавшись обозначениями, наиболее широко принятыми в литературе. Переобо-

Рис. 3.2. Расположение главных изоклин на фазовом портрете вольтерров- ской системы хищник жертва при разном соотношении параметров: а - о» б -
С I С2 С2
1, 3 - неустойчивые, 2 - устойчивая особая точка; в -
1, 2, 3 - неустойчивые, 4 - устойчивая особая точка значим
Система хищник-жертва в этих обозначениях имеет вид:

Свойства решений системы (3.3.6) будем исследовать на фазовой плоскости N
1
ON
2
Система имеет два стационарных решения. Их легко определить, приравняв нулю правые части системы. Получим:

Отсюда стационарные решения:

Рассмотрим подробнее второе решение. Найдем первый интеграл системы (3.3.6), не содержащий t. Умножим первое уравнение на -72, второе - на -71 и результаты сложим. Получим:
Теперь разделим первое уравнение на N и умножим на € 2, а второе разделим на JV 2 и умножим на е. Результаты снова сложим:
Сравнивая (3.3.7) и (3.3.8), будем иметь:

Интегрируя, получим:

Это и есть искомый первый интеграл. Таким образом, система (3.3.6) является консервативной, поскольку имеет первый интеграл движения, величину, представляющую собой функцию переменных системы N и N 2 и не зависящую от времени. Это свойство позволяет конструировать для вольтерровских систем систему понятий, аналогичную статистической механике (см. гл. 5), где существенную роль играет величина энергии системы, неизменная во времени.
При каждом фиксированном с > 0 (что соответствует определенным начальным данным) интегралу соответствует определенная траектория на плоскости N 1 ON 2 , служащая траекторией системы (3.3.6).
Рассмотрим графический способ построения траектории, предложенный самим Вольтерра. Заметим, что правая часть формулы (3.3.9) зависит только от Д г 2, а левая - только от N. Обозначим
Из (3.3.9) следует, что между X и Y имеется пропорциональная зависимость
На рис. 3.3 изображены первые квадранты четырех систем координат XOY, NOY , N 2 OX и Д Г 10N 2 так, чтобы все они имели общее начало координат.
В левом верхнем углу (квадрант NOY) построен график функции (3.3.8), в правом нижнем (квадрант N 2 OX) - график функции Y. Первая функция имеет min при Ni = а вторая - max при N 2 = ?-
Наконец, в квадранте XOY построим прямую (3.3.12) для некоторого фиксированного С.
Отметим точку N на оси ON . Этой точке соответствует определенное значение Y(N 1), которое легко найти, проведя перпендикуляр

Рис. 3.3.
через N до пересечения с кривой (3.3.10) (см. рис. 3.3). В свою очередь, значению К(Д^) соответствует некоторая точка М на прямой Y = сХ и, следовательно, некоторое значение X(N) = Y(N)/c, которое можно найти, проведя перпендикуляры AM и MD. Найденному значению (эта точка отмечена на рисунке буквой D) соответствуют две точки Р и G на кривой (3.3.11). По этим точкам, проводя перпендикуляры, найдем сразу две точки Е" и Е ", лежащие на кривой (3.3.9). Их координаты:

Проводя перпендикуляр AM , мы пересекли кривую (3.3.10) еще в одной точке В. Этой точке соответствуют те же Р и Q на кривой (3.3.11) и те же N и Щ. Координату N этой точки можно найти, опустив перпендикуляр из В на ось ON. Таким образом, мы получим точки F" и F", также лежащие на кривой (3.3.9).
Исходя из другой точки N, тем же самым образом получим новую четверку точек, лежащих на кривой (3.3.9). Исключение составит точка Ni = ?2/72- Исходя из нее, получим только две точки: К и L. Это будут нижняя и верхняя точки кривой (3.3.9).
Можно исходить не из значений N , а из значений N 2 . Направляясь от N 2 к кривой (3.3.11), поднимаясь затем до прямой У = сХ, а оттуда пересекая кривую (3.3.10), также найдем четыре точки кривой (3.3.9). Исключение составит точка No = ?1/71- Исходя из нее, получим только две точки: G и К. Это будут самая левая и самая правая точки кривой (3.3.9). Задавая разные N и N 2 и получив достаточно много точек, соединив их, приближенно построим кривую (3.3.9).
Из построения видно, что эго замкнутая кривая, содержащая внутри себя точку 12 = (?2/721 ?1/71)» исходящая из определенных начальных данных N ю и N20. Взяв другое значение С, т.е. другие начальные данные, получим другую замкнутую кривую, не пересекающую первую и также содержащую точку (?2/721 ?1/71)1 внутри себя. Таким образом, семейство траекторий (3.3.9) есть семейство замкнутых линий, окружающих точку 12 (см. рис. 3.3). Исследуем тип устойчивости этой особой точки, воспользовавшись методом Ляпунова.
Так как все параметры е 1, ?2, 71,72 положительны, точка (N[расположена в положительном квадранте фазовой плоскости. Линеаризация системы вблизи этой точки дает:

Здесь n(t) и 7i2(N1, N 2 :
Характеристическое уравнение системы (3.3.13):

Корни этого уравнения чисто мнимые:

Таким образом, исследование системы показывает, что траектории вблизи особой точки представлены концентрическими эллипсами, а сама особая точка - центр (рис. 3.4). Рассматриваемая модель Вольтерра и вдали от особой точки имеет замкнутые траектории, хотя форма этих траекторий уже отличается от эллипсоидальной. Поведение переменных Ni, N 2 во времени показано на рис. 3.5.

Рис. 3.4.

Рис. 3.5. Зависимость численности жертвы N i и хищника N 2 от времени
Особая точка типа центр устойчива, но не асимптотически. Покажем на данном примере, в чем это заключается. Пусть колебания Ni(t) и ЛГгМ происходят таким образом, что изображающая точка движется по фазовой плоскости по траектории 1 (см. рис. 3.4). В момент, когда точка находится в положении М, в систему извне добавляется некоторое количество особей N 2, такое, что изображающая точка переходит скачком из точки М в точку Л/". После этого, если система снова предоставлена самой себе, колебания Ni и N 2 уже будут происходить с большими амплитудами, чем прежде, и изображающая точка двигается по траектории 2. Это и означает, что колебания в системе неустойчивы: они навсегда изменяют свои характеристики при внешнем воздействии. В дальнейшем мы рассмотрим модели, описывающие устойчивые колебательные режимы, и покажем, что на фазовой плоскости такие асимптотические устойчивые периодические движения изображаются при помощи предельных циклов.
На рис. 3.6 изображены экспериментальные кривые - колебания численности пушных зверей в Канаде (по данным компании Гудзонова залива). Эти кривые построены на основании данных по числу заготовленных шкурок. Периоды колебаний численности зайцев (жертв) и рысей (хищников) примерно одинаковы и порядка 9 10 лет. При этом максимум численности зайцев опережает, как правило, максимум численности рысей на один год.
Форма этих экспериментальных кривых значительно менее правильная, чем теоретических. Однако в данном случае достаточно того, что модель обеспечивает совпадение наиболее существенных характеристик теоретических и экспериментальных кривых, г.е. величин амплитуды и сдвига фаз между колебаниями численностей хищников и жертв. Гораздо более серьезным недостатком модели Вольтерра является неустойчивость решений системы уравнений. Действительно, как уже говорилось выше, любое случайное изменение численности того или другого вида должно привести, следуя модели, к изменению амплитуды колебаний обоих видов. Естественно, что в природных условиях животные подвергаются бесчисленному количеству таких случайных воздействий. Как видно из экспериментальных кривых, амплитуда колебаний численностей видов мало изменяется от года к году.
Модель Вольтерра - эталонная (базовая) для математической экологии в той же мере, в какой модель гармонического осциллятора является базовой для классической и квантовой механики. При помощи этой модели на основе очень упрощенных представлений о характере закономерностей, описывающих поведение системы, сугубо математи-
Глава 3

Рис. 3.6. Кинетические кривые численности пушных зверей по данным пуш ной компании Гудзонова залива (Сетон-Томсон, 1987) ческими средствами было выведено заключение о качественном характере поведения такой системы - о наличии в такой системе колебаний численности популяции. Без построения математической модели и ее использования такой вывод был бы невозможен.
В рассмотренном нами выше самом простом виде системе Воль- терра присущи два принципиальных и взаимосвязанных недостатка. Их «устранению» посвящена обширная эколого-математическая литература. Во-первых, включение в модель любых, сколь угодно малых, дополнительных факторов качественным образом меняет поведение системы. Второй «биологический» недостаток модели заключается в том, что в нее не включены принципиальные свойства, присущие любой паре взаимодействующих по принципу хищник-жертва популяций: эффект насыщения хищника, ограниченность ресурсов хищника и жертвы даже при избытке жертвы, возможность минимальной численности жертв, доступных для хищника, и пр.
С целью устранения этих недостатков были предложены разными авторами различные модификации системы Вольтерра. Наиболее ин- тересные из них будут рассмотрены в разделе 3.5. Здесь остановимся лишь на модели, учитывающей самоограничения в росте обеих популяций. На примере этой модели наглядно видно, как может меняться характер решений при изменении параметров системы.
Итак, рассматривается система

Система (3.3.15) отличается от ранее рассмотренной системы (3.3.6) наличием в правых частях уравнений членов вида -7uNf,
Эти члены отражают тот факт, что численность популяции жертв не может расти до бесконечности даже в отсутствие хищников в силу ограниченности пищевых ресурсов, ограниченности ареала существования. Такие же «самоограничения» накладываются и на популяцию хищников.
Для нахождения стационарных численностей видов iVi и N 2 приравняем к нулю правые части уравнений системы (3.3.15). Решения с нулевыми значениями численностей хищников или жертв не будут нас сейчас интересовать. Поэтому рассмотрим систему алгебраических
уравнений
 Ее решение
Ее решение
дает нам координаты особой точки. На параметры системы здесь следует положить условие положительности стационарных численностей: N > 0 и N 2 > 0. Корни характеристического уравнения системы, линеаризованной в окрестности особой точки (3.3.16):

Из выражения для характеристических чисел видно, что если выполнено условие
то численности хищников и жертв совершают во времени затухающие колебания, система имеет ненулевую особую точку устойчивый фокус. Фазовый портрет такой системы изображен на рис. 3.7 а.
Допустим, что параметры в неравенстве (3.3.17) так изменяют свои значения, что условие (3.3.17) обращается в равенство. Тогда характеристические числа системы (3.3.15) равны, а ее особая точка будет лежать на границе между областями устойчивых фокусов и узлов. При изменении знака неравенства (3.3.17) на обратный особая точка становится устойчивым узлом. Фазовый портрет системы для этот случая представлен на рис. 3.76.
Как и в случае одной популяции, для модели (3.3.6) можно разработать стохастическую модель, но для нее нельзя получить решение в явном виде. Поэтому мы ограничимся общими рассуждениями. Допустим, например, что точка равновесия находится на некотором расстоянии от каждой из осей. Тогда для фазовых траекторий, на которых значения JVj, N 2 остаются достаточно большими, вполне удовлетворительной будет детерминистическая модель. Но если в некоторой точке


Рис. 3.7. Фазовый портрет системы (3.3.15): а - при выполнении соотношения (3.3.17) между параметрами; б - при выполнении обратного соотношения между параметрами
фазовой траектории какая-либо переменная не очень велика, то существенное значение могут приобрести случайные флуктуации. Они приводят к тому, что изображающая точка переместится на одну из осей, что означает вымирание соответствующего вида. Таким образом, стохастическая модель оказывается неустойчивой, так как стохастический «дрейф» рано или поздно приводит к вымиранию одного из видов. В такого рода модели хищник в конечном счете вымирает, это может произойти либо случайно, либо вследствие того, что сначала элиминируется популяция его жертвы. Стохастическая модель системы хищник- жертва хорошо объясняет эксперименты Гаузе (Гаузе, 1934; 2000), в которых инфузория Paramettum candatum служила жертвой для другой инфузории Didinium nasatum - хищника. Ожидавшиеся согласно детерминистическим уравнениям (3.3.6) равновесные численности в этих экспериментах составляли примерно всего но пять особей каждого вида, так что нет ничего удивительного в том, что в каждом повторном эксперименте довольно быстро вымирали либо хищники, либо жертвы (а за ними и хищники).
Итак, анализ вольтерровских моделей взаимодействия видов показывает, что, несмотря на большое разнообразие типов поведения таких систем, незатухающих колебаний численности в модели конкурирующих видов не может быть вовсе. В модели хищник жертва незатухающие колебания появляются вследствие выбора специальной формы уравнений модели (3.3.6). При этом модель становится негрубой, что свидетельствует об отсутствии в такой системе механизмов, стремящихся сохранить ее состояние. Однако в природе и эксперименте такие колебания наблюдаются. Необходимость их теоретического объяснения послужила одной из причин для формулировки модельных описаний в более общем виде. Рассмотрению таких обобщенных моделей посвящен раздел 3.5.
Еще в 20-х гг. А. Лотка, а несколько позднее независимо от него В.Вольтера предложили математические модели, описывающие сопряженные колебания численности хищника и жертвы.
Модель состоит из двух компонентов:
С – численность хищников; N – численность жертв;
Предположим, что в отсутствии хищников популяция жертвы будет расти экспоненциально: dN/dt = rN. Но жертвы уничтожаются хищниками со скоростью, которая определяется частотой встреч хищника с жертвой, а частота встреч возрастает по мере увеличения численности хищника (С) и жертвы (N). Точное число встреченных и успешно съеденных жертв будет зависеть от эффективности, с которой хищник находит и ловит жертву, т.е. от а’ – «эффективности поиска» или «частоты нападений». Т.о., частота «успешных» встреч хищника с жертвой и, следовательно, скорость поедания жертв будет равна а’СN и в целом: dN/dt = rN – a’CN (1*).
В отсутствии пищи отдельные особи хищника теряют в весе, голодают и гибнут. Предположим, что в рассматриваемой модели численность популяции хищника в отсутствии пищи вследствие голодания будет уменьшаться экспоненциально: dC/dt = - qC, где q – смертность. Гибель компенсируется рождением новых особей со скоростью, которая, как полагают в этой модели, зависит от двух обстоятельств:
1) скорости потребления пищи, a’CN;
2)эффективности (f), с которой эта пища переходит в потомство хищника.
Т.о., рождаемость хищника равна fa’CN и в целом: dC/dt = fa’CN – qC (2*). Уравнения 1* и 2* составляют модель Лотки-Вольтера. Свойства этой модели можно исследовать, построить изоклины линии, соответствующие постоянной численности популяции, с помощью таких изоклин определяют поведение взаимодействующих популяций хищник-жертва.
В случае с популяцией жертвы: dN/dt = 0, rN = a’CN, или C = r/a’. Т.к. r и a’ = const, изоклиной для жертвы будет линия, для которой величина С является постоянной:
При низкой плотности хищника (С), численность жертвы (N) возрастает, наоборот – снижается.
Точно также для хищников (уравнение 2*) при dC/dt = 0, fa’CN = qC, или N = q/fa’, т.е. изоклиной для хищника будет линия, вдоль которой N постоянна: При высокой плотности жертвы, численность популяции хищника растет, а при низкой – снижается.
Их численность претерпевает неограниченные сопряженные колебания. Когда велико число жертв, численность хищников растет, что приводит к увеличению пресса хищников на популяцию жертвы и тем самым к уменьшению ее численности. Это снижение в свою очередь ведет к ограничению хищников в пище и падению их численности, которая вызывает ослабление пресса хищников и увеличение численности жертвы, что снова приводит к росту популяции хищников и т.д.
Популяции неограниченно долго совершают один и тот же цикл колебаний до тех пор, пока какое-либо внешнее воздействие не изменит их численность, после чего популяции совершают новые циклы неограниченных колебаний. На самом деле среда постоянно меняется, и численность популяций будет постоянно смещаться на новый уровень. Чтобы циклы колебаний, которые совершает популяция, были регулярными, они должны быть стабильными: если внешнее воздействие изменяет уровень численности популяций, то они должны стремиться к первоначальному циклу. Такие циклы называются устойчивыми, предельными циклами.
Модель Лотки-Вольтера позволяет показать основную тенденцию в отношениях хищник-жертва, которая выражается в возникновении колебаний численности в популяции жертвы, сопровождающихся колебаниями численности в популяции хищника. Основным механизмом таких колебаний является запаздывание во времени, свойственное последовательности состояния от высокой численности жертв к высокой численности хищников, затем к низкой численности жертв и низкой численности хищников, к высокой численности жертв и т.д.
5) ПОПУЛЯЦИОННЫЕ СТРАТЕГИИ ХИЩНИКА И ЖЕРТВЫ
Взаимоотношения «хищник - жертва» представляют звенья процесса передачи вещества и энергии от фитофагов к зоофагам или от хищников низшего порядка к хищникам высшего порядка. По характеру этих отношений различают три варианта хищников :
а) собиратели . Хищник собирает мелких достаточно многочисленных подвижных жертв. Такой вариант хищничества характерен для многих видов птиц (ржанок, зябликов, коньков и др.), которые затрачивают энергию только на поиск жертв;
б) истинные хищники . Хищник преследует и убивает жертву;
в) пастбищники . Эти хищники используют жертву многократно, например, оводы или слепни.
Стратегия добывания пищи у хищников направлена на обеспечение энергетической эффективности питания: затраты энергии на добывание пищи должны быть меньше энергии, получаемой при ее усвоении.
Истинные хищники делятся на
«жнецов», которые питаются обильными ресурсами (н, планктонные рыбы и даже усатый кит), и «охотников», которые добывают менее обильный корм. В свою очередь
«охотники» делятся на «засадников», подстерегающих добычу (например, щука, ястреб, кошка, жук- богомол), «искателей» (насекомоядные птицы) и «преследователей». Для последней группы поиск пищи не требует больших затрат энергии, но ее нужно много, чтобы овладеть жертвой (львы в саваннах). Впрочем, некоторые хищники могут сочетать элементы стратегии разных вариантов охоты.
Как и при отношениях «фитофаг - растение», ситуация, при которой все жертвы будут съедены хищниками, что в конечном итоге приведет и к их гибели, в природе не наблюдается. Экологическое равновесие между хищниками и жертвами поддерживается специальными механизмами , снижающими риск полного истребления жертв. Так, жертвы могут :
Убегать от хищника. В этом случае в результате адаптаций повышается подвижность и жертв, и хищников, что особенно характерно для степных животных, которым негде прятаться от преследователей;
Приобретать защитную окраску («притворяться» листьями или сучками) или, напротив, яркий цвет, Н.: красный, предупреждающий хищника о горьком вкусе. Общеизвестно изменение окраски зайца в разные времена года, что позволяет ему маскироваться летом в траве, а зимой на фоне белого снега. Адаптивное изменение окраски может происходить в разных стадиях онтогенеза: детеныши тюленей белые (цвет снега), а взрослые особи черные (цвет скалистого побережья);
Распространяться группами, что делает их поиск и промысел для хищника более энергоемким;
Прятаться в укрытия;
Переходить к мерам активной обороны (травоядные, имеющие рога, колючие рыбы), иногда совместной (овцебыки могут занимать «круговую оборону» от волков и т.д.).
В свою очередь хищники развивают не только способность к быстрому преследованию жертв, но и обоняние, позволяющее по запаху определить место нахождения жертвы. Многие виды хищников разрывают норы своих жертв (лисы, волки).
В то же время сами они делают все возможное для того, чтобы не обнаружить своего присутствия. Этим объясняется чистоплотность мелких кошачьих, которые для устранения запаха много времени тратят на туалет и закапывают экскременты. Хищники одевают «маскировочные халаты» (полосатость щук и окуней, делающих их менее заметными в зарослях макрофитов, полосатость тигров и т.д.).
Полной защиты от хищников всех особей в популяциях животных-жертв также не происходит, так как это привело бы не только к гибели голодающих хищников, но в конечном итоге - к катастрофе популяций жертв. В то же время при отсутствии или снижении плотности популяции хищников ухудшается генофонд популяции жертв (сохраняются больные и старые животные) и ввиду резкого увеличения их численности подрывается кормовая база.
По этой причине эффект зависимости численности популяций жертв и хищников - пульсация численности популяции жертвы, за которой с некоторым запаздыванием пульсирует численность популяции хищника («эффект Лотки - Вольтерры») - наблюдается редко.
Между биомассами хищников и жертв устанавливается достаточно устойчивое соотношение. Так, Р. Риклефс приводит данные о том, что соотношение биомасс хищника и жертвы колеблется в пределах 1:150 - 1:300. В разных экосистемах умеренной зоны США на одного волка приходится 300 мелких белохвостых оленей (вес 60 кг), 100 крупных оленей вапити (вес 300 кг) или 30 лосей (вес 350). Такая же закономерность выявлена в саваннах.
При интенсивной эксплуатации популяций фитофагов человек нередко исключает из экосистем хищников (в Великобритании, к примеру, есть косули и олени, но нет волков; в искусственных водоемах, где разводят карпа и другую прудовую рыбу, нет щук). В этом случае роль хищника выполняет сам человек, изымая часть особей популяции фитофага.
Особый вариант хищничества наблюдается у растений и грибов. В царстве растений встречается порядка 500 видов, способных ловить насекомых и частично их переваривать с помощью протеолитических ферментов. Хищные грибы образуют ловчие аппараты в виде маленьких овальных или шаровидных головок, расположенных на коротких веточках мицелия. Однако самый распространенный вид ловушки - клейкие трехмерные сети, состоящие из большого числа колец, образующихся в результате ветвления гиф. Хищные грибы могут ловить достаточно крупных животных, например, круглых червей. После того, как червь запутается в гифах, они прорастают внутрь тела животного и быстро его заполняют.
1.Постоянный и благоприятный уровень температуры и влажности.
2.Обилие пищи.
3.Защита от неблагоприятных факторов.
4.Агрессивный химический состав среды обитания (пищеварительные соки).
1.Наличие двух сред обитания: среда первого порядка - организм хозяина, среда второго порядка - внешняя среда.
Часто представители одного вида (популяции) питаются представителями другого вида.
Модель Лотки – Вольтерры – модель взаимного существования двух популяций типа «хищник – жертва».
Впервые модель «хищник – жертва» была получена А. Лоткой в 1925 году, который использовал ее для описания динамики взаимодействующих биологических популяций. В 1926 году независимо от Лотки аналогичные (к тому же более сложные) модели были разработаны итальянским математиком В. Вольтерра, глубокие исследования которого в области экологических проблем заложили фундамент математической теории биологических сообществ или т. н. математической экологии.
В математической форме предложенная система уравнений имеет вид:
где x – количество жертв, y – количество хищников, t – время, α, β, γ, δ – коэффициенты, которые отражают взаимодействия между популяциями.
Постановка задачи
Рассмотрим закрытое пространство, в котором существуют две популяции – травоядные («жертвы») и хищники. Считается, что животных не ввозят и не вывозят и что еды для травоядных животных достаточно. Тогда уравнение изменения числа жертв (только жертв) примет вид:
где $α$ – коэффициент рождаемости жертв,
$x$– размер популяции жертв,
$\frac{dx}{dt}$ – скорость прироста популяции жертв.
Когда хищники не охотятся, они могут вымирать, значит, уравнение для количества хищников (только хищников) примет вид:
Где $γ$ – коэффициент убыли хищников,
$y$ – размер популяции хищников,
$\frac{dy}{dt}$ – скорость прироста популяции хищников.
При встрече хищников и жертв (частота встреч прямо пропорциональна произведению) хищники уничтожают жертв с коэффициентом, сытые хищники могут воспроизводить потомство с коэффициентом. Таким образом, система уравнений модели примет вид:
Решение задачи
Построим математическую модель совместного существования двух биологических популяций типа «хищник – жертва».
Пусть две биологические популяции совместно обитают в изолированной среде. Среда является стационарной и обеспечивает в неограниченном количестве всем необходимым для жизни один из видов – жертвы. Другой вид – хищник – также обитает в стационарных условиях, но питается только жертвами. В роли хищников могут выступать коты, волки, щуки, лисы, а в роли жертв – куры, зайцы, караси, мыши соответственно.
Для определенности рассмотрим в роли хищников – котов, а в роли жертв – кур.
Итак, куры и коты живут в некотором изолированном пространстве – хозяйственном дворе. Среда предоставляет курам питание в неограниченном количестве, а коты питаются только курами. Обозначим через
$х$ – количество кур,
$у$ – количество котов.
Со временем количество кур и котов меняется, но будем считать $х$ и $у$ непрерывными функциями от времени t. Назовем пару чисел $х, у)$ состоянием модели.
Найдем каким образом изменяется состояние модели $(х, у).$
Рассмотрим $\frac{dx}{dt}$ – скорость изменения количества кур.
Если котов нет, то количество кур возрастает и тем быстрее, чем больше кур. Будем считать зависимость линейной:
$\frac{dx}{dt} a_1 x$,
$a_1$ – коэффициент, который зависит только от условий жизни кур, их естественной смертности и рождаемости.
$\frac{dy}{dt}$ – скорость изменения количества котов (если нет кур), зависит от количества котов y.
Если кур нет, то количество котов уменьшается (у них нет пищи) и они вымирают. Будем считать зависимость линейной:
$\frac{dy}{dt} - a_2 y$.
В экосистеме скорость изменения количества каждого вида также будем считать пропорциональным его количеству, но только с коэффициентом, зависящим от количества особей другого вида. Так, для кур этот коэффициент уменьшается с увеличением количества котов, а для котов возрастает с увеличением количества кур. Будем считать зависимость также линейной. Тогда получим систему дифференциальных уравнений:
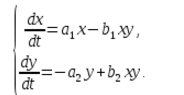
Данная система уравнений называется моделью Вольтерра-Лотки.
a1, a2, b1, b2 – числовые коэффициенты, которые называют параметрами модели.
Как видно, характер изменения состояния модели (x, y) определяется значениями параметров. Изменяя данные параметры и решая систему уравнений модели, можно исследовать закономерности изменения состояния экологической системы.
С помощью программы MATLAB система уравнений Лотки-Вольтерра решается следующим образом:
На рис. 1 представлено решение системы. В зависимости от начальных условий решения разные, чему отвечают разные цвета траекторий.
На рис. 2 представлены те же решения, но с учетом оси времени t (т.е. наблюдается зависимость от времени).


